Bild 2.2
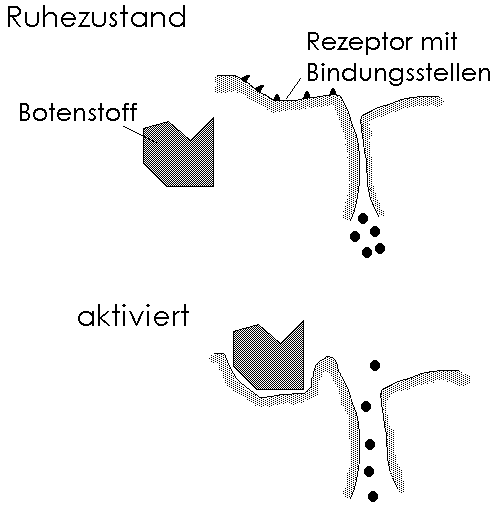
Ein Botenstoff löst eine Wirkung an einem Rezeptor aus: Der Botenstoff tritt mit den Bindungsstellen eines Rezeptors in Kontakt. Hierdurch verformt sich das Rezeptorprotein, so daß ein Ionenkanal eröffnet wird.
Computersysteme - Ebenbilder der Natur?
Claudia Borchard-Tuch, Springer
, ISBN 3-540-67042-4(siehe auch
Herausforderung: Biocomputer)Kapitel 2
Zelle und Computer
2.1 Die Zelle
Nahezu alle Lebensvorgänge vom Stoffwechsel bis zur Vererbung können als Informationsprozesse aufgefaßt werden, deren Wurzeln bis auf die Ebene der biologischen Makromoleküle zurückreichen. Für den Informationsaustausch zwischen Lebewesen sind nicht einmal Sinnesorgane, ein Gehirn oder ein Nervensystem notwendig. Auch Zellen selbst verfügen über ein kompliziertes System der Informationsverarbeitung und -weitergabe.
Die zelluläre Informationsübertragung ist ein biochemischer Vorgang. Moleküle verschlüsseln Reize aus der Umwelt, die gemäß der inneren Zellbiologie umgewandelt und so in Stoffwechseländerungen, Verhaltensweisen und extrazelluläre Reize übersetzt werden. Eine einzelne Zelle beherbergt ein kompliziertes chemisches Netzwerk, in dem die Signale durch direkten Kontakt zwischen Molekülen übermittelt werden. Hierbei ist - anders als im Computer - jede Einheit mit jeder anderen verbunden.
2.1.1 Die Umwelt
|
Die Zelle ist ein offenes System, das mit seiner Umwelt ständig über einen Stoff-, Energie- und Informationsaustausch verbunden ist. Zahlreiche Umwelteinflüsse wie beispielsweise Licht, Temperatur, Wassergehalt oder chemische Substanzen bestimmen das äußere Milieu einer Umweltsignal ¯ |
|
Genaktivierung ¯ |
|
Proteinsynthese ¯ |
|
Transmittersynthese ¯ |
|
physiologische Wirkungen und Verhalten |
Bild 2.1
Umweltreize verändern über eine Kette molekularer Umwandlungen Physiologie und VerhaltenZelle und lösen zellphysiologische und generell biochemische Informationsprozesse aus.
Zum einen verändern Umweltreize die Art und Weise, in der die in den Genen enthaltene Information in bestimmte Eigenschaften einer Zelle oder eines Organismus übersetzt wird, d.h. die Genexpression. Damit haben Umweltreize Einfluß auf die genetisch gesteuerte Synthese funktionell bedeutsamer Enzyme, die ihrerseits Stoffwechselprodukte mit physiologischer Wirkung entstehen lassen.
Neben der Genexpression verändern Umweltreize die Struktur von bereits produzierten Genprodukten, etwa Enzymen oder Gerüsteiweißen (sogenanntes posttranslationales Processing). Je nach Zelltyp werden die Genprodukte chemisch verändert, beispielsweise acetyliert, amidiert, glykosyliert, methyliert oder phosphoryliert.
In dieser Weise stehen einzelne Zellen in engem Kontakt mit ihrer Umwelt und kommunizieren mit ihr. In mehrzelligen Organismen ist der Kommunikationsbedarf noch erheblich höher. Vielfältige Aufgaben sind auf zahlreiche Zellen, Gewebe und Organe verteilt, die weit voneinander entfernt liegen können. Um ihre Funktionen aufeinander abzustimmen, müssen sie untereinander gezielt Botschaften über weite Entfernungen weitergeben. Dies geschieht über das Hormon- und das Nervensystem. In beiden Systemen verständigen sich die Zellen mit Hilfe kommunikativer Substanzen. Zu ihnen gehören die von Nerven freigesetzten Überträgerstoffe, "Transmitter", sowie die aus endokrinen Drüsen (z.B. Schilddrüse, Nebenniere, Bauchspeicheldrüse) abgegebenen Hormone, die die Zelle über den Blutweg oder die die Zellen umgebende Flüssigkeit erreichen.
Obwohl kommunikative Substanzen auf den ersten Blick nur der Informationsübertragung innerhalb eines Organismus zu dienen scheinen, ist bei ihnen die Umwelt nicht ohne Einfluß und modifiziert ihre Konzentrationen im Körper. So läßt eine streßreiche Umgebung den Spiegel zahlreicher Hormone steigen, wie z.B. den des Adrenalins oder den der Schilddrüsenhormone.
2.1.2 Der Informationsaustausch zwischen Zellen
Zellen übermitteln einander Informationen, indem sie Substanzen ausschütten, die als chemische Boten wirken. Zumeist dienen Rezeptoren den Botenstoffen als Anlegestelle. Der Begriff des Rezeptors ist mehrdeutig. Zum einen bezeichnet er eine Sinneszelle (z.B. den Photorezeptor des Gesichtssinnes); zum anderen - in unserem Sinne - jede Struktur, die eine andere, einen sogenannten Botenstoff, bindet und so eine Information überträgt und eine Wirkung auslöst.
Rezeptoren sind große Aggregate aus vielen Molekülen, deren besondere Verknüpfung jeder Rezeptorart eine charakteristische Struktur verleiht. Durch die Anpassung der Rezeptor-Bindungsstelle an die Struktur des gebundenen Botenstoffs (das Schlüssel-Schloß-Prinzip) kommt es zu einer Verformung des Rezeptormoleküls, wodurch eine Wirkung ausgelöst wird.
Das Schlüssel-Schloß-Prinzip ist sehr wichtig für eine sichere Übertragung der im Botenstoff gespeicherten Information an den richtigen Empfänger. Da ein Botenstoff der äußeren Umgebung jede Stelle der Zelloberfläche erreichen kann, muß er erkennen, welche Zellstruktur seine Botschaft empfangen und beeinflußt werden soll und welche nicht. Diese Unterscheidungsfähigkeit macht eine große Strukturvielfalt rezeptorgebundener Moleküle notwendig: Jede Mo-
|
|
Bild 2.2 Ein Botenstoff löst eine Wirkung an einem Rezeptor aus: Der Botenstoff tritt mit den Bindungsstellen eines Rezeptors in Kontakt. Hierdurch verformt sich das Rezeptorprotein, so daß ein Ionenkanal eröffnet wird. |
lekülart löst an einem zu ihr passenden Rezeptor für sie spezifische Wirkungen aus.
Ebenso wie der Botenstoff den zu ihm passenden Rezeptor findet, ist auch eine einzelne rezeptortragende Zellstruktur fähig, zwischen den verschiedenen Botenstoffen, die sie zu binden vermag, zu unterscheiden. Je nach Art des Botenstoffs nimmt sie eine bestimmte Information auf und richtet ihr Verhalten danach aus. Kontraktilität, Sekretion und Zellteilung werden so beeinflußt, die Genexpression ermöglicht und die Immunabwehr.
Einzeller ohne Nervensystem erkennen mit Hilfe von Rezeptoren zahlreiche chemische Stoffe. Chemotaktische Bakterien, die durch gelöste Stoffe in der Umgebung angezogen oder abgestoßen werden, finden selbständig chemische Nahrungsquellen oder fliehen vor Giften, da sie in ihrer Zellmembran über Rezeptoren verfügen, die Moleküle aus der Zellumgebung an sich binden.
Neben Proteinen sind zumeist auch Kohlenhydrate am Aufbau eines Rezeptors beteiligt. So sind die an der Oberfläche von Zellmembranen lokalisierten Kohlenhydrate Bestandteile zellspezifischer Glykoproteinrezeptoren. Sie dienen zum einen der Anheftung infektiöser Bakterien und Viren sowie verschiedener Toxine, Hormone und anderer Moleküle. Zum anderen vermitteln sie während der Embryonalentwicklung die Wanderung von Zellen. Eine besonders wichtige Rolle spielen Kohlenhydrate bei Wechselwirkungen zwischen einzelnen Zellen, wie z.B. der Zell-Zell-Erkennung.
Zellen werden von einer Membran begrenzt, die den Zellinhalt gegen die Umgebung abgrenzt und ein weitgehend eigenständiges Innenleben ermöglicht. Sie besteht aus einer doppelten Protein-Phospholipidschicht. Jedes Substrat, jeder Wirkstoff, der das Zellinnere erreichen soll, jedes Stoffwechselprodukt, das die Zelle passiert, muß die Membran überwinden. Sie wirkt bei diesen Prozessen manchmal als Barriere, indem sie bestimmten Substanzen den Durchtritt erschwert oder unmöglich macht.
Zum Teil transportiert sie auch Stoffe, die die Membran spontan nur schwer passieren würden. Zum kontrollierten Austausch mit der Umgebung dienen in die Membran eingebettete Proteine und Glykoproteine, seien es energieverbrauchende Pumpen (z.B. Na
+/K+-ATPase),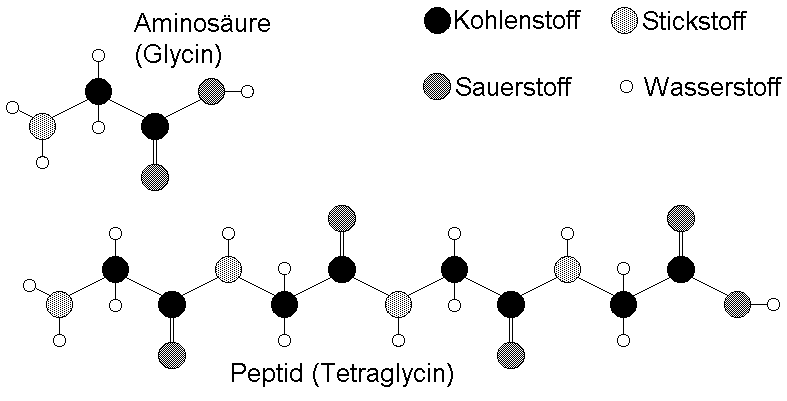
Bild 2.3
Vier Aminosäuren des Typs Glycin lassen sich nur auf eine einzige Art miteinander verknüpfen - zu dem Peptid Tetraglycin.andere Transportproteine (carrier, z.B. Na
+/Glucose-Cotransport) und Ionenkanäle (Na+-Kanal, Ca++-Kanal).Zwei Zellen erkennen einander über komplementäre Strukturen an der Außenseite ihrer Membranen: Die betreffende Struktur auf der einen Zelle enthält eine codierte biologische Information, die von der Struktur auf der anderen verstanden werden kann.
Sind Proteine an der Zellerkennung beteiligt, werden kurze Abfolgen von Aminosäuren - Peptide - erkannt. Manche Anheftungsprozesse laufen beispielsweise über sogenannte Integrine; das sind Oberflächenproteine, die passende Peptide erkennen.
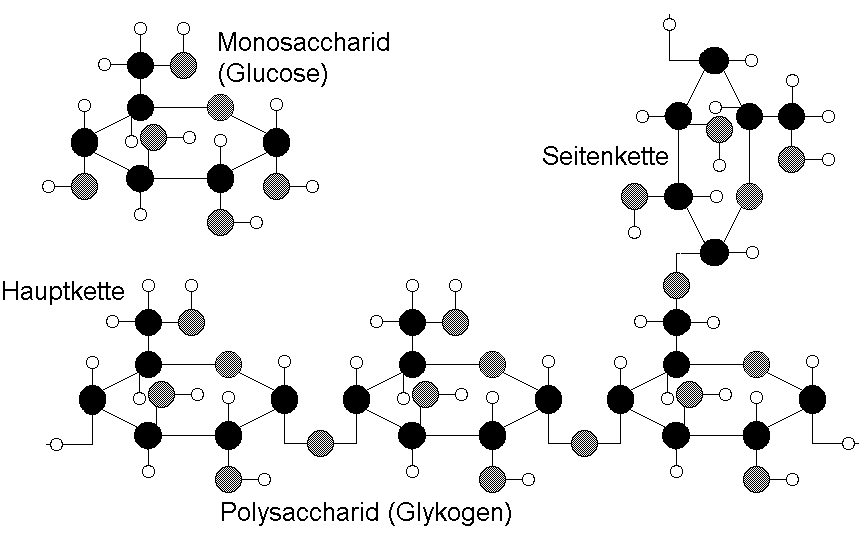
Bild 2.4
Monosaccharide (Einfachzucker) können auf mannigfaltige Art miteinander verknüpft werden. Glykogen ist ein ausschließlich aus Glucose aufgebauter Vielfachzucker (Polysaccharid) mit stark verzweigter Struktur. Nach drei bis fünf Glucosemolekülen erfolgt eine Verzweigung der linearen Bindung. Glykogen ist nur eine der zahlreichen Möglichkeiten, Glucosemoleküle miteinander zu verbinden.Bausteine von Proteinen lassen sich nur auf eine Art linear verknüpfen. Da es von vier verschiedenen Nukleinsäuren bzw. Aminosäuren 4! = 24 unterschiedliche Anordnungsmöglichkeiten (Permutationen) gibt, lassen sie sich zu 24 verschiedenen Tetranukleotiden oder Tetrapeptiden zusammenfügen.
Im Vergleich zu Proteinen verfügen Kohlenhydrate über komplexere Verknüpfungsmöglichkeiten. Zwei identische Monosaccharide lassen sich bereits auf elf unterschiedliche Arten miteinander verknüpfen. Aus wenigen verschiedenen Disacchariden kann so eine unvorstellbare Vielfalt verzweigter und unverzweigter Kohlenhydrate entstehen: Vier verschiedene Einfachzucker können zu 35 560 unterschiedlichen Tetrasacchariden verknüpft werden.
Ihre Strukturvielfalt macht Zuckerpolymere zu wichtigen Informationsträgern; sie vermögen weit mehr Informationen pro Gewichtseinheit zu speichern als Nukleinsäuren und Proteine. So können Monosaccharide als Buchstaben einer Schrift verstanden werden, deren Worte aus Verknüpfungen von Einfachzuckern bestehen.
2.1.3 Kommunikationsmoleküle
Höhere mehrzellige Organismen verfügen über ein Hormon- und ein Nervensystem, in dem sich die Zellen mit Hilfe kommunikativer Substanzen, d.h. mit Hilfe von Hormonen bzw. Transmittern, verständigen.
Eine Nervenzelle sendet einzelne Signale jeweils an eine spezifische Gruppe von Zielzellen: Muskelzellen, Drüsenzellen oder andere Nervenzellen. Sie übermittelt ihre Botschaft, indem sie einen Transmitter von speziellen Stellen aus, den Synapsen, zur Zielzelle schickt. So wird das Nervensystem beispielsweise von einem dichten Netz von Nervenzellen durchsetzt, deren Neurotransmitter Noradrenalin ist. Nahezu alle zerebralen Funktionen lassen sich durch eine Beeinflussung aktivierender noradrenerger Mechanismen regulieren - Aufmerksamkeit und Konzentration, Schlaf-Wach-Rhythmus, Nahrungsaufnahme und Temperaturregulation. Hierbei sind Hormon- und Nervensystem eng miteinander verbunden. So sind Noradrenalin enthaltende Nervenenden im Hypothalamus (Teil des Zwischenhirns, vgl. 5.2) an der Regulation von Hormonausschüttungen beteiligt.
Im Vergleich zu Neurotransmittern wirken Hormone gewöhnlich weniger direkt. Zwar gibt es die sogenannten exokrinen Drüsen, die ihr Sekret über ein Gangsystem abgeben und damit direkt ihren Wirkort erreichen, der zumeist eine Oberfläche ist. Doch findet die häufigste Form der hormonellen Kommunikation im endokrinen, also innersekretorischen, System statt: Endokrine Drüsen haben keinen Ausführungsgang, sie geben ihre Hormone an die Blutbahn ab. Diese Hormone werden über den Blutweg transportiert und erreichen so die Organe, an denen sie ihre Wirkungen entfalten. Zu den wichtigsten endokrinen Drüsen zählen die Hirnanhangsdrüse (an der Unterseite des Gehirns), die Schilddrüse und die Nebenschilddrüsen (im Halsbereich), die Bauchspeicheldrüse und die Nebennieren.
Ihre physiologischen Wirkungen entfalten Nerven- und Hormonsystem in ähnlicher Weise: Beide lassen ihre Kommunikationsmoleküle in Kontakt mit spezifischen Rezeptoren einer Zielzelle treten.
Auf diese Weise ist gewährleistet, daß die durch das Hormon oder den Transmitter zu übermittelnde Information die richtigen Empfängerzellen erreicht - eine außerordentlich wichtige Funktion: Da alle mit dem Blut zirkulierenden Hormone jedes Körperorgan erreichen, muß ein Hormon erkennen, welche Organzellen es beeinflussen soll und welche nicht.
Hormonrezeptoren finden sich nicht nur auf der äußeren Zellmembran, sondern auch innerhalb der Zelle. Steroidhormone dringen beispielsweise durch die Zellhaut und finden im Zellinneren ein spezifisches zytoplasmatisches Bindungsprotein.
Die durch Hormone und Neurotransmitter hervorgerufenen Wirkungen in mehrzelligen Lebewesen entsprechen den durch Umweltreizen verursachten in einzelligen Organismen. Aber Umweltreize spielen auch in mehrzelligen Organismen eine wichtige Rolle:
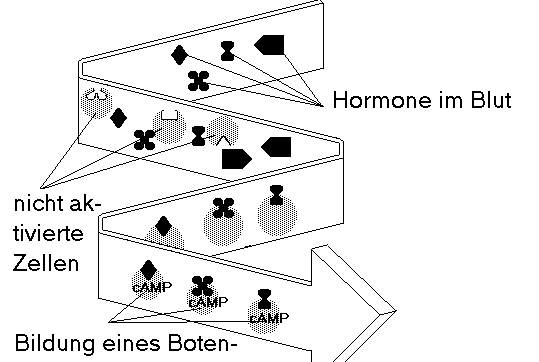

Bild 2.5
Jedes Hormon wirkt spezifisch nur auf Zellen eines bestimmten Zielgewebes. Das Hormon paßt zu seiner Bestimmungszelle wie ein Schlüssel zu einem Schloß. Gehen Hormon und Zielzelle eine Verbindung ein, so bilden sich Botenstoffe der zweiten Stufe (vgl. 2.1.4) - in diesem Fall cAMP (modifiziert nach Benner).Indem sie Transmitter und Hormone kontrollieren, haben sie Zugriff auf die intrazellulären informationsverarbeitenden Systeme. Da die meisten Zellen mehrere Arten von Kommunikationsmolekülen synthetisieren, verfügen sie über ein enormes Potential an Signalkombinationen. Auf diese Weise bildet ein Spektrum aus Transmittern und Hormonen die Wirklichkeit ab; Erfahrungen werden in Form einzigartiger Kombinationen von molekularen Symbolen verschlüsselt.
Eine der schnellsten Reaktionen auf plötzliche körperliche oder seelische Belastungen - Streß - ist beispielsweise die Ausschüttung von Adrenalin aus der Nebenniere. Über eine Stimulierung sogenannter Adrenozeptoren kommt es zu einer Reihe intrazellulärer Folgereaktionen. Innerhalb von Sekunden nach Adrenalinausschüttung finden im Körper zahlreiche sichtbare Veränderungen statt. Der Organismus wird zu höheren Leistungen befähigt und seine Energiereserven werden mobilisiert - so wie es in Situationen des Kampfes oder der Flucht notwendig ist: Das Blut wird in die Muskeln umgeleitet, wo es benötigt wird; die Lungen erweitern sich zur vermehrten Sauerstoffaufnahme; die Muskeln werden vorgespannt, um plötzliche und rasche Bewegungen zu gestalten; Glucose und Fett werden von der Leber ins Blut abgegeben. Das Herz schlägt
|
Die Nebennieren geben Adrenalin an das Blut ab. |
|
Die Pupillen erweitern sich: Das Sehfeld wird größer. |
|
Hautgefäße verengen sich, so daß das Blut in die Muskeln fließt. |
|
Das Herz schlägt schneller und kraftvoller |
|
Die Bronchien erweitern sich, so daß die Lunge mehr Sauerstoff aufnehmen kann. |
|
Die Leber gibt Zucker und Fette als Energielieferanten an das Blut ab. |
Bild 2.6
Antworten auf Stressschneller, und der Blutdruck steigt, so daß sich der Energieverbrauch erhöht. Der Mensch ängstigt oder ärgert sich.
Hormon- und Transmitterwirkungen infolge von Umwelt-reizen, die nur Sekunden bis Minuten einwirken, sind oft noch tagelang nachweisbar. So läßt uns der Streß auch dann nicht zur Ruhe kommen, wenn er längst vorüber ist. Nach einem hektischen Tag zirkuliert Adrenalin in erhöhter Konzentration in unserem Körper und hält ihn im Zustand höchster Alarmbereitschaft - obwohl es Zeit zum Schlafen ist.
Das Beispiel zeigt, daß (auch unerwünschte) Hormonwirkungen gespeichert werden und zur Gedächtnisbildung beitragen. Im allgemeinen speichern Zellen Gedächtnis als biochemische Veränderung oder Veränderung der Struktur. Zellsysteme speichern Wissen in Verbindungen ihrer Zellen, d.h. als Verknüpfungsmuster. Auf diese Weise wird die Information eingebettet in ein Netz von Assoziationen. Ein Beispiel ist die metaplastische Umwandlung von Zellen einer Gewebeart in die einer anderen Form. Hierbei erwerben Zellen neues Wissen, das sie dauerhaft speichern. So kann am unverhornten Plattenepithel der Mundschleimhaut als Folge einer chronischen chemischen oder mechanischen Reizeinwirkung die Fähigkeit zur Hornbildung erworben werden - es resultieren Stellen verhornten Plattenepithels.
Jedoch verfügen Zellen noch über andere Gedächtnisformen. So ist das Wissen über das, was körpereigen und körperfremd ist, im Immunsystem festgelegt (vgl. Kapitel 3), und genetisches Wissen ist in den Erbanlagen gespeichert (vgl. Kapitel 4).
2.1.4 Die Informationsverarbeitung einer Zelle
Einem Rezeptor kommt eine duale Funktion zu: Indem er einen molekularen Boten bindet, erkennt er ein Signal. Zum anderen leitet er Signale weiter bzw. verarbeitet sie und löst damit Effekte aus. Hierbei können - je nach Art der gebundenen Substanz - entgegengesetzte Wirkungen ausgelöst werden. Bindet ein Rezeptor einen Agonisten, geht er in seine aktive Konformation über, und die Rezeptorwirkung wird verstärkt. Bindet der Rezeptor dagegen einen Antagonisten, so begünstigt dies die Ausbildung der inaktiven Konformation, und die Rezeptorwirkung wird geschwächt.
Rezeptoren kommen innerhalb und außerhalb einer Zelle vor. Intrazellulär gelegene Zellen binden ein Botenmolekül und werden so aktiviert. Der Rezeptor-Botenstoff-Komplex dringt sodann in den Zellkern ein und reagiert dort mit DNA. Im folgenden wird m-RNA synthetisiert, die nach Ausschleusung aus dem Zellkern die Bildung eines spezifischen Proteins hervorruft, das dann die eigentliche Wirkung auslöst.
Auf der äußeren Zellmembran sitzende Rezeptoren können einfach-membrangängig sein. Binden sie ein Botenmolekül, löst dies am entgegengesetzten Rezeptorende eine Phosphorylierung aus, worauf der Rezeptor-Botenstoff-Komplex von der Membran ins Zellinnere transportiert wird. Auf diese Weise erfolgt z.B. die Aufnahme bestimmter Eiweiß-Fett-Verbindungen (Low-density-Lipo-proteine) in die Zelle.
Ein Ionenkanal-Rezeptor bildet durch die Zellmembran einen für Ionen durchgängigen Kanal. Bindet der Rezeptor einen Botenstoff, so wird der Kanal entweder geöffnet oder geschlossen. Die Folge ist ein verstärkter oder verringerter Austausch der entsprechenden Ionen.
Hat ein Botenstoff einen G-Protein-gekoppelten Rezeptor gebunden, wird ein Guanin-Nukleotide-bindendes, membran-verankertes Kopplungsprotein, ein sogenanntes G-Protein, angeregt, eine Reaktionskaskade innerhalb der Zelle in Gang zu setzen. G-Proteinen an der Innenseite von Zellmembranen kommt eine wesentliche Vermittlerrolle bei der Signalübertragung innerhalb der Zelle vor. Zunächst aktivieren sie effektorische Enzyme, die als erste Stufe der zellulären Reaktion die Bildung weiterer Botenstoffe innerhalb der Zelle fördern.
Bereits bei einfachen einzelligen Organismen sorgen effektorische Enzyme für die Anpassung des Verhaltens an die Anforderungen der Umgebung und die damit verbundene Aufnahme und Verar-
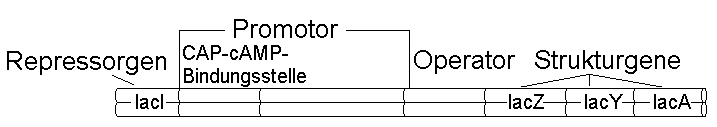
Bild 2.7
Schematische Darstellung des lac-Operons von Escherichia coli mit dem unmittelbar anschließenden lac-Repressor-Gen. Das Operon besteht aus drei Strukturgenen (lacZ, lacY und lacA) und einer Promotor-Operator-Sequenz. Das Gen lacI definiert den lac-Repressor. Verbindet dieser sich mit dem Operator, wird die Übersetzung der genetischen Information des lac-Operons unmöglich gemacht.beitung von Information. So verfügt das Darmbakterium Escherichia coli über eine besondere Fähigkeit: Es kann Lactose (Milchzucker) als einzige Kohlenstoff- und Energiequelle nutzen, da es ein Enzym besitzt, das Lactose in Glucose (Traubenzucker) und Galactose (4-stereoisomere Form der Glucose) zerlegt - die ß-Galactosidase. Dieses Enzym wird nur dann in größeren Mengen hergestellt, wenn dies erforderlich ist, d.h. wenn die Zellen auf einem Nährmedium heranwachsen, das keine Glucose, sondern Lactose enthält. Das ß-Galactosidase-codierende Gen (lacZ) ist ein "induzierbares Gen", das nur dann zur Synthese der ß-Galactosidase führt, wenn der Zelle genügend ß-Galactoside zur Verfügung stehen.
Damit es zur Ausbildung des ß-Galactosidase-codierenden Gens kommt, ist ein positives Signal erforderlich. Dieses besteht aus einem Komplex aus zyklischem AMP (cAMP) und einem Protein (genannt CAP, catabolite activator protein). Der Komplex geht mit dem Gen eine Bindung ein und aktiviert es. Über mehrere Zwischenschritte führt dies schließlich zu einer vermehrten Bildung der ß-Galactosidase.
ß-Galactosidase ist nicht das einzige Enzym, dessen Herstellung sich nur bei ausreichender ß-Galactosidzufuhr lohnt. Auch die ß-Galactosid-Permease (codiert durch Gen lacY) und die ß-Galactosid-Transacetylase (codiert durch Gen lacA) wandeln Galactoside um. Daher wendet die Natur ihren Sparmechanismus gleichzeitig auf alle drei Enzyme an: Die Gene lacZ, lacY und lacA liegen auf der DNA benachbart und werden zusammen aktiviert. Gemeinsam mit einer sogenannten Promotor-Operator-Sequenz, d.h. einem bestimmten, für die Steuerung der gesamten Gengruppe verantwortlichen DNA-Abschnitt, bilden sie ein Operon (vgl. 4.4.3).
Neben cAMP sind andere intrazelluläre Botenstoffe bekannt. cGMP (zyklisches Guanosinmonophosphat) entsteht nach Aktivierung des Enzyms Guanylatcyclase. Verändert ein Reiz das Potential
|
|
extrazelluläre Signale |
|
|
|
|
erste Boten: |
Neurotransmitter, Hormone |
|
|
ß |
|
|
intrazelluläre Signale |
|
|
|
|
zweite Boten: |
cAMP, cGMP, Ca2+, Phosphytidylinosi-tolsystem |
|
|
ß |
|
|
cAMP-abhängige Proteinkinase cGMP-abhängige Proteinkinase Ca2+-abhängige Proteinkinase |
|
|
ß |
|
dritte Boten: |
Substrate der Proteinkinasen |
|
|
|
|
Vierte...nte Boten: |
ß |
|
|
ß |
|
|
ß |
|
|
physiologische Antworten |
Bild 2.8
Beispiele für Signalkaskaden. Extrazelluläre Signale lösen über eine Serie von intrazellulären Transduktionen physiologische Antworten aus.der Zellhülle, so kann es auch zu einem Einstrom von Calcium durch spannungsgesteuerte Ionenkanäle in die Zelle kommen.
Calcium sowie cAMP und cGMP wandeln Enzyme der zweiten Stufe, die man als Kinasen bezeichnet, aus unwirksamen Formen in wirksame um. Die aktivierten Kinasen übertragen Phosphatgruppen auf Enzyme der dritten Stufe. Dieser Vorgang der Phosphorylierung ist eine in allen Zellen benutzte Methode, um den chemischen Zustand von Molekülen zu ändern. Phosphorylierte Enzyme sind enzymatisch aktiv, das heißt, sie wirken wie Katalysatoren, die eine chemische Reaktion in Gang bringen und beschleunigen können. Die aktivierten Enzyme bringen dann die Veränderungen hervor, die der äußere Reiz auslösen soll.
So beeinflussen sie beispielsweise in verschiedenen Bakterienarten die Drehrichtung der Geißel und damit die Schwimmrichtung des Bakteriums. Das Bakterium bewegt sich dorthin, wo es mehr und bessere Nahrung findet und entfernt sich aus Gebieten, wo es Hunger leidet und wo Gefahren drohen. Ein solches Verhalten entspricht den Reflexen im Tierreich. Da die Signalübertragung vom Rezeptor bis zur Geißel nicht länger als eine Zehntelsekunde dauert, ist die Reaktion des Bakteriums ausreichend schnell.
Phosphatidyl-4,5-diphosphat (PIP2)
Proteinkinase
Diacylglycerol Inositol-1,4,5-triphosphat (IP3)
Calcium
Bild 2.9
PhosphatidylinositolkaskadeAndere Umweltreize aktivieren eine weitere intrazelluläre Reaktionskette, die sogenannte Phosphatidylinositolkaskade. Diese läßt intrazelluläre Phospholipidsignale entstehen, die ebenfalls Enzymaktivitäten regulieren. Ein Enzym, die Phospholipase C, ist dabei von besonderem Interesse. Sie baut ein Phospholipid in der Zellmembran gleich zu zwei zweiten Botenstoffen ab, nämlich Diacylglycerol und Inositol-1,4,5-triphosphat (IP3), das aus Speichern im Zellinneren einen weiteren Botenstoff - Calcium - freisetzt.
Phosphorylierungs- und Phosphatidylinositolkaskade beeinflussen sich gegenseitig: Information fließt von einer Reaktionskette zur anderen, woraus sich komplizierte erregende und hemmende Wechselwirkungen ergeben.
2.1.5 Schlußfolgerung
Räumliche Struktur und biologische Organisation einer Zelle legen fest, in welcher Weise Umweltreize in Funktionsänderungen übersetzt werden. Hierbei haben Umweltreize Zugriff auf jene intrazellulären Vorgänge, die den Kern der zellulären Informationsverarbeitung darstellen. Reize lösen biologische Mechanismen aus, und biologische Regeln bestimmen die Reizumwandlung. Indem Botenstoffe spezifisch Umweltreize in zelluläre Informationen umwandeln, haben sie Übersetzerfunktion. Hierbei hängt der Gehalt der übersetzten zellulären Information von den Operationsregeln und der Arbeitsweise des Zellsystems ab - die Bedeutung eines Moleküls ist reiz- und systemspezifisch. Sie ergibt sich u.a. aus der Rolle des Moleküls in einem komplexen biochemisch-verhaltensphysiologischen Netz - das Molekül im Reagenzglas verliert seinen Informationsgehalt.
2.2 Der Automat
Die Zelle ist ein höchst komplexes Gebilde. Indem wir eine Modellvorstellung von ihr entwickeln, nehmen wir ihr ihre Komplexität und ahmen die Informationsverarbeitung natürlicher Zellsysteme in vereinfachter Weise nach. Es entsteht ein Abbild der Wirklichkeit, ein Ding, das uns auch im alltäglichen Leben häufig begegnet und das nach eigenen Gesetzen selbständig wirkt - ein sogenannter endlicher Automat. Die Reduktion eröffnet zum einen die Möglichkeit, Zellstrukturen mit Hilfe von Computern zu simulieren. Zum anderen lassen sich durch endliche Automaten all die Elemente nachbilden, die die Grundlagen heutiger Digitalrechner bilden.
2.2.1 Was ist ein endlicher Automat?
Ein Automat ist eine Maschine, die sich selbst steuert. Sie reagiert auf bestimmte Ereignisse mit ausgewählten Funktionen, ohne daß ein Mensch einzugreifen braucht. Ein Eingabesignal bewirkt, daß sie in einen anderen Zustand übergeht. Ein endlicher Automat kann sich nur in einer endlichen Zahl verschiedener Zustände befinden.
Überall begegnen uns in unserem täglichen Leben endliche Automaten. Kaffeeautomat, Schreibmaschine, Musikinstrument, Taschenrechner - sie alle sind Beispiele. Ihnen ist gemeinsam, daß sie auf einige Signale und Eingaben der Umwelt in bestimmter Weise reagieren.
2.2.2 Die Informationsaufnahme eines endlichen Automaten
Umweltsignale können z.B. dadurch erzeugt werden, daß Tasten gedrückt werden wie bei Schreibmaschine, Taschenrechner oder Klavier, Münzen eingeworfen werden wie bei Fahrkarten- und Spielautomat oder Zeichen eingelesen werden, die evtl. zuvor auf einem Speichermedium wie z.B. Magnetplatten oder Bändern gespeichert waren.
|
Start Wasserbehälter leer kein Filterpapier eingelegt kein Kaffeepulver eingefüllt Strom abgeschaltet |
|||||
|
¯ F |
¯ W |
||||
|
Wasserbehälter leer Filterpapier eingelegt kein Kaffeepulver eingefüllt Strom abgeschaltet |
Wasserbehälter gefüllt kein Filterpapier eingelegt kein Kaffeepulver eingefüllt Strom abgeschaltet |
||||
|
|
¯ W |
¯ F |
|
||
|
K |
Wasserbehälter gefüllt Filterpapier eingelegt kein Kaffeepulver eingefüllt Strom abgeschaltet |
|
|||
|
|
|
||||
|
Wasserbehälter leer Filterpapier eingelegt Kaffeepulver eingefüllt Strom abgeschaltet |
K |
||||
|
¯ W |
|
||||
|
Wasserbehälter gefüllt Filterpapier eingelegt Kaffeepulver eingefüllt Strom abgeschaltet |
|||||
|
¯ S |
|||||
|
Erfolgszustand Wasserbehälter gefüllt Filterpapier eingelegt Kaffeepulver eingefüllt Strom eingeschaltet |
|||||
Bild 2.10
Die Zustände eines KaffeeautomatenFür jeden Automaten gibt es somit eine für ihn typische Menge von elementaren Signalen, mit denen die Umwelt auf ihn einwirken kann und die der Automat "versteht". Diese Menge bezeichnet man als Eingabealphabet des Automaten.
Nehmen wir als Beispiel einen Kaffeeautomaten, so gibt es einzelne Funktionen, die durch Eingabesignale gesteuert werden, nämlich Wasser einfüllen, Filterpapier einlegen, Kaffeepulver einfüllen und Strom einschalten. Kürzen wir diese elementaren Operationen durch W, F, K und S ab, dann sind zum Beispiel WFKS oderauch FKWS und FWKS als korrekte Eingabefolgen anzusehen, die nach einiger Zeit zum gewünschten Ergebnis, nämlich heißem Kaffee, führen.
Dagegen sind die Folgen WFS (gefiltertes, heißes Wasser), KWS (Kaffeesatz in der Kanne) oder SFK (Überhitzung des Gerätes) weit weniger erfolgreich.
|
2.2.3 |
Die Informationsverarbeitung eines endlichen Automaten |
Durch einzelne Eingaben und somit auch Eingabefolgen wird der "innere Zustand" eines Automaten verändert. Der Automat kann verschiedene endlich viele Zustände annehmen, die diskret, d.h. klar voneinander abgrenzbar und einzeln benennbar, sein müssen. Zu jedem Zustand können Ein- und Ausgaben gehören. Beim Übergang zwischen den Zuständen wird der Modellcharakter des endlichen Automaten besonders deutlich: Zustandswechsel erfolgen ohne Zeitverbrauch. Diese blitzschnellen Zustandswechsel können nur in der Vorstellung existieren. Stellt man sich beispielsweise den Wechsel zwischen Bewegung und Stillstand ohne Zeitverbrauch vor, sind beide diskrete Zustände.
Bei einer korrekten Bedienungsfolge, z.B. WFKS, wird der Anfangszustand des Kaffeeautomaten (Wasserbehälter leer, kein Filterpapier eingelegt, kein Kaffeepulver eingefüllt, Strom abgeschaltet) überführt in den Erfolgszustand (Wasserbehälter gefüllt, Filterpapier eingelegt, Kaffeepulver im Filter, Strom eingeschaltet), wobei einige Zwischenzustände durchlaufen werden.
Ein solcher Zwischenzustand ist zum Beispiel (Wasserbehälter gefüllt, Filterpapier eingelegt, noch kein Kaffeepulver eingefüllt, Strom abgeschaltet), der durch die Eingabe K in den nächsten Zwischenzustand (Wasserbehälter gefüllt, Filterpapier eingelegt, Kaffeepulver im Filter, Strom abgeschaltet) überführt wird. Der Automat reagiert also auf eine Eingabe durch Übergang in einen anderen Zustand, und zwar in Abhängigkeit vom momentanen inneren Zustand und vom gerade erhaltenen Eingabezeichen.
Zustände sowie Ein- und Ausgaben legen die Maschine fest. Es gibt keine zusätzlichen Informationen und insbesondere keine Möglichkeit, Informationen zu speichern.
2.2.4 Zelluläre Automaten
Die Erfinder der zellulären Automaten, John von Neumann und Stanislaw Ulam, wollten ein Modell sich wechselseitig beeinflussender Teilkomponenten entwerfen, mit dessen Hilfe die Entwicklung vernetzter Systeme studiert werden konnte. Als Vorbild für ihren Entwurf sahen sie die natürlichen Zell- und Nervenzellverbände. So sind die Teilkomponenten ihres Systems sich selbst reproduzierende Automaten, d.h. Maschinen, die einen anderen, völlig gleichen Automaten erzeugen können. Diese Automaten sind in einem gleichförmigen Gitter angeordnet, von dem jedes Feld einen endlichen Automaten verkörpert. Die identischen Automaten, auch "Zellen" genannt, stehen untereinander in enger Verbindung: Jeder erhält Eingabeinformationen von benachbarten Automaten, wobei die Nachbarschaftsverbindung in der gesamten Anordnung einheitlich ist. Der Zustand einer Automatenzelle kann durch wenige Zahlenwerte beschrieben werden, und alle Zellen ändern ihren Zustand entsprechend den gleichen Regeln und zur gleichen Zeit.
Mit Hilfe zellulärer Automaten lassen sich die unterschiedlichsten Systeme sich lokal beeinflussender Teile modellieren - von der Nebelbank bis zum Weltraum. Zelluläre Automaten können helfen, Antworten auf die unterschiedlichsten Fragen zu finden, z.B.: Wie arbeiten Computer in einem bestimmten Netzwerk zusammen?
Der Entwurf von John von Neumann und Stanislaw Ulam hatte natürliche Zellsysteme zum Vorbild, und zelluläre Automaten können auch als Modell biologischer, zur Selbstreproduktion fähiger Zellsysteme angesehen werden. Hierbei wird jedoch der Modellcharakter zellulärer Automaten besonders deutlich. Jede ihrer Zellen ist von einer recht eintönigen Umwelt umgeben: Die Zelle empfängt eine kleine Menge diskreter Eingabesignale, die sich durch einfache Zahlenwerte darstellen lassen und der unmittelbaren Nachbarschaft entstammen. Es gibt keine Rezeptoren und keine Kommunikationsmoleküle, die einen differenzierten Informationsaustausch auch zwischen weiter entfernten Strukturen regeln. Die komplizierte Signalkaskade der wirklichen Zelle vereinfacht sich so zu einem blitzartigen Wechsel zwischen diskreten Zuständen.
|
2.2.5 |
Die Informationsverarbeitung eines zellulären Automaten |
In Abhängigkeit von voreingestellten Signalen schaltet jede Automatenzelle zwischen verschiedenen Zuständen um. Hierbei bestimmt die Zelle ihren neuen Wert aus ihrem vorherigen und den alten Inhalten ihrer Nachbarzellen. Der Zustand einer Automatenzelle kann durch wenige Zustandswerte beschrieben werden - oft sind dies nur die Werte 0 und 1. Darin ähnelt sie einem Digitalcomputer, in dem jede Information ebenfalls nur durch digitale Einheiten - der begrenzten Abfolge unterschiedlicher Bits - ausgedrückt wird.
Auf den ersten Blick erscheinen die Entwicklungsmöglich-keiten eines zellulären Automaten recht eingeschränkt, doch dieser Schein trügt: Viele einfache Teile tun sich zusammen, um ein komplexes Ganzes zu formen.
Auch wenn die einzelne Zelle kaum Möglichkeiten der Entscheidung hat, ist die Kombinationsbreite für die Zustände des gesamten zellulären Automaten gewaltig. Sie hängt allein ab von der Zahl aller Zellen des Zellraumes. Enthält der Zellraum eines Automaten N Zellen, die nur zwei Zustände annehmen können, so gibt es 2
N verschiedene Möglichkeiten, die zwei verschiedenen Zustandswerte auf alle Zellen zu verteilen.Das ganze Spektrum der Entwicklungsmöglichkeiten hängt sowohl von der Zahl der Zellzustände als auch von der Größe der Nachbarschaft ab. Jede mögliche Kombination der Zustandswerte einer Nachbarschaft kann mit einem eigenen neuen Zustand der Zelle verknüpft werden.
2.2.6 Nachbarschaftliche Beziehungen
Alle Zellen sind von gleicher Gestalt. Sie können die Form eines Quadrates, eines Sechsecks oder eines Dreiecks annehmen. Am häufigsten findet man ein zweidimensionales rechteckiges Gitter, in dem die Zellen wie die Felder eines Schachbrettes angeordnet sind.
Bild 2.11
Zweidimensionale Zellräume: rechteckige, hexagonale und dreieckige Gitterstruktur|
|
Bild 2.12 Von Neumann- (links) und Moore-Nachbarschaft (rechts) |
Die geometrische Grundform einer Zelle legt ihre nachbarschaftlichen Beziehungen fest: Hat eine Zelle in einem Sechseck-Gitter sechs Zellen in ihrer unmittelbaren räumlichen Nachbarschaft, so sieht dies in einem rechteckigen Gitter ganz anders aus. Hier ist eine Zelle direkt benachbart zu vier anderen Zellen (sogenannte von-Neumann-Nachbarschaft). Statt sich nur auf diese vier Nachbarn einer Zelle zu beschränken, können auch die vier an den Ecken angrenzenden Zellen als weitere Nachbarn hinzugenommmen werden (Moore-Nachbarschaft).
In eindimensionalen Automaten liegen die Variationsmöglichkeiten im Radius der Nachbarschaft. Einen Effekt auf die Entwicklung einer Zelle müssen nicht nur die zwei unmittelbar angrenzenden Zellen haben - auch über eine Entfernung von zwei oder drei Zellen kann die Entwicklung beeinflußt werden.
Welchen Einfluß der Zustand der Nachbarn auf die eigene Entwicklung nimmt, also die Art der Wechselwirkungen, legt die Regel des Automaten fest. Sie beschreibt das Programm, das für alle Zellen gleich ist und das wie in einem Parallelrechner von ihnen simultan ausgeführt wird: In jedem Zeittakt werden die Werte aller Zellen gleichmäßig nach einer vorgegebenen Regel verändert.
2.2.7 Das Spiel des Lebens
Im Spiel des Lebens, 1968 von John Horton Conway erfunden, entwickeln und vermehren sich die Akteure selbständig wie Lebewesen. Es sind die Elemente eines Zellularautomaten, die über ein zweidimensionales rechteckiges Feld verteilt sind. Jedes dieser Elemente hat die Form eines Quadrates und steht in Moore-nachbarschaftlicher Beziehung zu seinen angrenzenden Zellen. Ein Element kann nur zwei Zustände annehmen, beschrieben durch 0 und 1. 1 erhält es, wenn sein eigener Zustand und der seiner Nachbarn zusammen 3 ergibt bzw. auch 4, wenn das Element selbst denWert 1 hatte. In allen anderen Fällen wird dem Element der Wert 0 zugeordnet.
Hinter diesen Bedingungen steckt die Absicht, die Entwicklung eines wirklichen Zellsystems zu modellieren. Ein Element repräsentiert eine lebende Zelle, die in eine zweidimensionale rechteckige Gitterwelt hineingeboren wird. Je nach Bevölkerungsdichte in ihrer Umgebung bleibt sie am Leben oder stirbt: Ein Zustandswert von 1 bedeutet, daß sie lebt; ein Wert von 0 dagegen entspricht ihrem Tod. Eine Zelle wird zum Leben erweckt, wenn es in den acht um sie herum liegenden Gitterplätzen eine ideale Bevölkerungsdichte von genau drei lebenden Zellen gibt. Auch das Überleben einer Zelle hängt von der Zahl ihrer Lebensgefährten in der Nachbarschaft ab. Streiten sich mehr als drei lebende Zellen um den lokalen Lebensraum in ihrer Nachbarschaft, stirbt die Zelle. Ein gleiches Schicksal ereilt sie aber auch, wenn sie nicht mindestens zwei lebende Nachbarn findet und an Einsamkeit zugrunde geht. Nur bei zwei oder drei lebenden Zellen in ihrer Umgebung sind somit die Voraussetzungen für das eigene Überleben gesichert.
Es ist nicht vorhersehbar, wie sich die Zellmuster im Spiel des Lebens entwickeln, und es entstehen überraschende und vielfältige Zellmuster. Einige Muster verschwinden und werden nie mehr gesehen - sie sterben aus. Andere Muster durchlaufen zyklische Konfigurationen, bei denen nach gewisser Zeit das Ausgangsmuster wieder erreicht wird. Manche Muster wandern über die Bildfläche in horizontaler, vertikaler oder diagonaler Richtung, und wieder andere Muster enden in einer gleichbleibenden Konfiguration (vgl. Bild 2.13).
|
|
Bild 2.13 Beispiele stationärer Muster, die sich aus vier lebenden Zellen entwickeln |
Das in diesen Automaten umgesetzte Prinzip lokaler Wechselwirkungen einfacher Bausteine erfaßt auf eine leicht zugängliche Weise Grundcharakteristika der sich selbstorganisierenden Welt natürlicher Zellsysteme. Man kann sich mit ihnen den Phänomenen der Komplexität annähern, ohne erst einen umfassenden theoretischen Apparat im Detail zu studieren.
Spiegeln Zellularautomaten zum einen das Verhalten na-türlicher Zellsysteme wider, ähnelt ihr Verhalten zum anderen auch dem von Computern. Mit den Regeln des Spiel des Lebens lassen sich all die Elemente nachbauen, die die Grundlagen heutiger Digitalrechner bilden. Es können beliebige Symbole codiert, manipuliert und gespeichert werden, d.h. Informationen verarbeitet werden, - mit nichts anderem als den Elementen und Regeln des zellulären Spiels selbst.
|
2.2.8 |
Wie das Spiel des Lebens einen Computer nachbildet |
Sind es Moleküle, die in biologischen Systemen Informationen verarbeiten, so ist es in Computern elektrischer Strom. Die Ausführung einer Mikrooperation, d.h. einer in der Hardware eines Computers vorgesehenen, festverdrahteten Elementaroperation, bedeutet die Umwandlung einer Information in ein elektrisches Signal, d.h. in einen der beiden Zustände "Strom fließt" bzw. "Strom fließt nicht". In dieser elektrischen Form werden Informationen im Rechner verarbeitet, d.h. daß sie gespeichert oder arithmetisch bzw. logisch miteinander verknüpft werden. Bei arithmetischen Operationen verkörpern die beiden elektrischen Zustände (Strom fließt bzw. Strom fließt nicht) die binären Ziffern 1 und 0, während sie bei der Verknüpfung logischer Ausdrücke für wahr und falsch stehen. Elektrische Schaltkreise ermöglichen die Verarbeitung der eingegebenen Bitfolgen. Die Basis all dieser Schaltungen sind drei elementare Grundbausteine, die sogenannten logischen Gatter: die logischen
|
UND (Ù ) |
ODER ( Ú ) |
NICHT ( Ø ) |
![]()
|
UND (0,0) = 0 UND (0,1) = 0 UND (1,0) = 0 UND (1,1) = 1 |
ODER (0,0) = 0 ODER (0,1) = 1 ODER (1,0) = 1 ODER (1,1) = 1 |
NICHT (0) = 1 NICHT (1) = 0 |
Bild 2.14
Die drei logischen GatterVerknüpfungen UND, ODER und NICHT. Die UND-Funktion ergibt den Wert wahr, wenn alle verknüpften Aussagen wahr sind. Die ODER-Funktion liefert den Wert wahr, wenn mindestens eine der verknüpften Aussagen wahr ist. Die NICHT-Funktion verwandelt den Wahrheitswert einer Aussage in sein Gegenteil. Die gesamte Informationsverarbeitung eines Computers beruht auf dem Hintereinanderschalten dieser drei logischen Schaltungen. Dabei wird zunächst jede Aktion des Computers auf die Manipulation einzelner Bits zurückgeführt.
Informationsverarbeitung in Computern und Zellularautomaten entsprechen sich. Im Spiel des Lebens fließt natürlich kein Strom. Jede Information wird auf einem zweidimensionalen Gitter verarbeitet und durch "belebte" Gitterplätze dargestellt. Doch gibt es Zellkonfigurationen, die genau wie in wirklichen Computern jede beliebige Eingabe binär verschlüsseln können. Es sind die sogenannten Gleiter. Ein Gleiter ist eine bestimmte Anordnung "belebter" Zellen, die in vier aufeinanderfolgenden Generationen ihre Gestalt verändert, um dann nach genau vier Generationen in gleicher Form, nur diagonal um eine Zelle versetzt, wieder aufzutauchen. Ein Gleiter ist somit ein Muster, das sich in einem ewigen Lauf unverändert durch die Gitterwelt bewegt.
|
t = 0 |
t = 1 |
t = 2 |
t = 3 |
t = 4 |
|
|
|
Bild 2.15 Lebenszyklus eines Gleiters |
||||
Folgen von Gleitern sind im Spiel-des-Lebens-Computer das Gegenstück zu elektrischen Impulsen. Das Spiel des Lebens kennt eine Konfiguration, die regelmäßig in Abständen von 30 Zeitschritten einen Gleiter erzeugen kann, die sogenannte Gleiterkanone. Die Periodenlänge der Gleiterkanone, d.h. 30 Zeitschritte, bestimmt die Länge eines Arbeitstaktes des zellulären Computers. Ist ein Gleiter in einem Takt vorhanden, so ist dies gleichbedeutend zur "Strom an"-Phase im wirklichen Computer. Wird zu Beginn des nächsten Arbeitstaktes, d.h. nach 30 Zeitschritten, kein Gleiter auf die Reise geschickt, so ist der "Strom" ausgeschaltet. Da jede beliebige Folge von Gleitern möglich ist, kann genau wie in wirklichen Computern jede beliebige Eingabe codiert werden.
Auch die drei logischen Gatter können mit Hilfe von Gleitern erzeugt werden. Da sich beim Zusammenstoß zwei Gleiter gegenseitig vernichten können, kann so ein NICHT-Gatter nachgebildet werden, das die Eingabe 0 (falsch) in eine 1 (wahr) verwandelt und umgekehrt: Ist eine Folge von Gleitern Eingabestrom eines NICHT-Gatters, so wird eine zweite kontinuierliche Folge von Gleitern derart auf diesen Eingabestrom gerichtet, daß seine Gleiter (Einsen) in Nichtgleiter (Nullen) verwandelt werden. Falls der Eingabestrom an einer Stelle keinen Gleiter enthält, kann der Gleiter aus dem kontinuierlichen Strom unbehelligt passieren, so daß eine 0 in eine 1 umgewandelt wird.
Mit kollidierenden Gleiterströmen lassen sich auch die anderen Gattertypen, d.h. UND- und ODER-Gatter nachbilden, so daß sich auf der Grundlage des Spiels des Lebens ein Computer vollständig konstruieren läßt.
|
2.2.9 |
Wie ein Computer einen zellulären Automaten nachbildet |
Der vorangegangene Abschnitt zeigte, daß die Informations-verarbeitung eines zellulären Automaten in die eines Computers übersetzt werden kann. Umgekehrt ist es möglich, mit Hilfe eines Computers einen zellulären Automaten zu simulieren. So ist die Folge von Binärzellen im Speicher eines Computers mit den Anfangswerten eines Zellularautomaten vergleichbar. Während der Entwicklung eines Zellularautomaten wird diese in den Anfangswerten gespeicherte Information verarbeitet. Die Entwicklung aus einem Anfangszustand kann man als eine Berechnung ansehen, die die Informationen des jeweiligen Zustandes verarbeitet.
|
|
Bild 2.16 Torus |
Jeder der Entwicklungsschritte läßt sich mit Hilfe eines Computerprogramms simulieren, so daß das Ergebnis nach einer bestimmten Zeit gefunden werden kann. Das Programm legt zunächst einen Bereich für die Zellen fest, wobei die Zellen den Speicherelementen im Computer entsprechen. Idealerweise wäre dieser Bereich unendlich groß, aber dies ist praktisch unmöglich. Um das Problem des offenen Randes zu vermeiden, kann man die Ränder des Zellraums miteinander verbinden. Im eindimensionalen Raum wird so aus einem einfachen Streifen von Zellen ein Ring, in zwei Dimensionen ein Torus (vgl. Bild 2.16). Die zweite Möglichkeit der künstlichen Randerweiterung ist die Spiegelung der Randzellen.
Hierbei wird die nicht vollständige Nachbarschaft einfach am tatsächlichen Rand des Raums gespiegelt, so daß jede Randzelle auch hier genauso viele Nachbarn besitzt wie die Zelle im Innern.
Das Programm sucht nun einzeln nacheinander jede Zelle auf, untersucht deren Nachbarzellen und berechnet den Wert für den nächsten Zustand der Zelle. Hierbei muß der Inhalt einer Zelle unverändert bleiben, bis ihr Wert von all den Zellen geprüft worden ist, deren Nachbar sie ist.
Auf einem normalen Digitalrechner ausgeführt, durchläuft das Programm nacheinander die einzelnen Schritte. Dabei bildet es die Operationen vieler zusammengeschalteter, gleichzeitig laufender Computer nach, d.h. es simuliert sie.
Gibt es eine wirksamere Möglichkeit, an das Ergebnis zu gelangen? Gibt es eine Abkürzung der schrittweisen Simulation, ein Verfahren, das das Ergebnis vieler Entwicklungsschritte abgibt, ohne jeden Schritt durchlaufen zu haben? Der Rechner könnte so die Entwicklung des Zellularautomaten ohne ausdrückliche Simulation vorherbestimmen. Eine einfache Überlegung führt zu dem Schluß, daß es keine allgemeine Vereinfachung geben kann, die die Entwicklung eines beliebigen Zellularautomaten bestimmt: Die meisten Zellularautomaten verfügen über eine universelle Rechenfähigkeit, d.h. daß sie jedes berechenbare Problem lösen können. Würde ein Verfahren bekannt sein, das das Verhalten dieser Zellularautomaten schneller lieferte als sich der Automat selbst entwickelte, könnte man damit jede Berechnung beschleunigen. Da dies etwas Unmögliches ist (wie sich mathematisch beweisen läßt), folgt, daß sich die Entwicklung eines beliebigen Zellularautomaten nicht ohne weiteres vereinfachen läßt. Dieses Problem ist irreduzibel, nicht zu vereinfachen - das Ergebnis kann man nur durch schrittweise Simulation erreichen.
Allgemein scheint bei biologischen Systemen die rechnerische Irreduzibilität weit verbreitet zu sein: So könnte sich beispielsweise herausstellen, daß man die Entwicklung einer Zelle aus ihrem genetischen Code nur durch Verfolgung eines jeden einzelnen Entwicklungsschrittes bestimmen kann.
Zellularautomaten sind Modelle von Zellsystemen, deren Art der Informationsverarbeitung der eines Computers entspricht: Wie die vorangegangenen Abschnitte zeigten, sind zum einen Zellularautomaten in der Lage, Computer nachzuahmen und zum anderen ist es möglich, mit Hilfe von Computern Zellularautomaten zu simulieren.
Jedoch sind Zellularautomaten stark vereinfachte Modellvorstellungen von Zellsystemen, die wenige ausgewählte Eigenschaften der Wirklichkeit berücksichtigen. Bei genauer Betrachtung unterscheiden sich Zellen einerseits sowie Zellularautomaten und Computer andererseits deutlich in ihrer Art, Informationen zu verarbeiten.
2.3 Computer
2.3.1 Die Hardware
Ein Computer verfügt über Bauteile, die sich so zusammenschalten lassen, daß sie logische und arithmetische Operationen ausführen können. Dieses Bauteil ist der Transistor, ein elektronischer Schalter, der den Stromfluß regelt. Ein Transistor besteht aus drei halbleitenden Schichten. Die beiden äußeren Schichten werden Emitter und Kollektor genannt, die mittlere Schicht heißt Basis. Die drei Schichten sind in ihren elektrischen Eigenschaften so aufeinander abgestimmt, daß eine kleine Verschiebung des Basis-Kollektor-Stroms eine sehr viel größere Änderung des Emitter-Kollektor-Stroms hervorruft.
Ein hoher Kollektorstrom läßt sich als 1, ein niedriger als 0 interpretieren. Durch Kombination von Transistoren und anderen Bauelementen kann man UND-, ODER- und NICHT-Funktionen aufbauen (vgl. 2.2.8). Das UND-Glied liefert einen hohen Ausgangsstrom, falls alle Eingangsströme groß sind. Das ODER-Glied erzeugt einen hohen Ausgangsstrom, wenn mindestens einer der
|
Bild 2.17 Transistor |
Eingangsströme groß ist. Das NICHT-Glied liefert für einen großen Eingangsstrom einen kleinen Ausgangsstrom und umgekehrt.
Transistoren lassen sich miteinander zu beliebig komplizierten Schaltungen kombinieren. Diese Schaltungen nehmen wenig Platz in Anspruch: Auf einem sogenannten Chip, d.h. einem winzigen Plättchen aus Silizium, können auf wenigen Quadratmillimetern über 100 000 Transistoren eingebettet werden. Transistorschaltungen verbrauchen auch wenig Rechenzeit: Ein zentraler Taktgeber schaltet die Transistoren bis zu 1 Million mal pro Sekunde.
Daten sind im Rechner Bit für Bit gespeichert, und das Speicherelement für 1 Bit kann genau zwei Zustände festhalten. Zur kurzfristige Speicherung dienen zumeist Register, für die längerfristige Speicherung Magnetkernspeicher.
An vielen Stellen im Computer muß bei der Verarbeitung Information kurzfristig festgehalten werden, oder bestimmte Daten müssen immer wieder sofort greifbar sein. Dafür gibt es die Register, die für das Abspeichern und die Wiedergabe von Information nur einen Bruchteil der Zeit benötigen, die beim Arbeitsspeicher für Lesen und Schreiben notwendig ist. Ein Register besteht aus einzelnen Speicherelementen, von denen jedes 1 Bit speichern kann. Meistens benutzt man als Speicherelement eine aus Transistoren bestehende elektronische Schaltung, die als Flipflop bezeichnet wird. Die Information am Flipflopausgang kann jederzeit unmittelbar abgenommen werden - es bedarf zum Übernehmen des gespeicherten Inhalts keines besonderen Lesevorgangs.
Das Einschreiben einer Information in ein Register bezeichnet man als Laden des Registers. Die vor dem Laden im Register stehende Information wird durch einen kurzen Stromimpuls einfach überschrieben.
Ringförmige Kerne aus Ferrit sind die Speicherelemente des Kernspeichers. Fließt durch einen dieser Ferritkerne ein kurzer Stromstoß hindurch, so wird der Kern in eine bestimmte Richtung magnetisiert. Diesen magnetischen Zustand behält er so lange, bis er durch einen Stromstoß anderer Richtung in entgegengesetzter Richtung ummagnetisiert wird. Man legt fest, daß die Magnetisierung des Kerns in der einen Richtung einer gespeicherten 1, die entgegengesetzte Magnetisierung einer gespeicherten 0 entspricht. Jeder Kern kann somit genau 1 Bit speichern.
2.3.2 Zelle und Computer
Im Vergleich zum Computer verfügt eine Zelle über wesentlich differenziertere Möglichkeiten der Informationsverarbeitung. So könnten sämtliche Aufgaben eines Computers auch von einem Netzwerk chemischer Moleküle ausgeführt werden: Datenaufnahme, -integration und -verstärkung ebenso wie die Speicherung von Informationen und ihre digitale Verarbeitung mit Hilfe logischer Entscheidungen.
Es gibt Proteine, die wie ein digitaler Computer genau zwei Zustände einnehmen können: So entspricht etwa der aktive, phos-
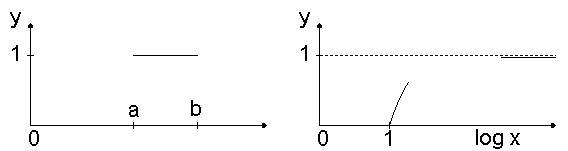
Bild 2.18
Fuzzy-Menge. Die Menge der klassischen Mengentheorie wird beschrieben durch eine Abbildung, die als charakteristische Funktion bezeichnet wird. Diese Abbildung ordnet jedem Element der Menge den Wert 1 und allen Elementen, die nicht zur Menge gehören, den Wert 0 zu. Für ein Intervall [a,b] der reellen Zahlen erhalten wir dann das Diagramm oben links.Eine unscharfe Menge oder fuzzy-Menge einer gegebenen Grundmenge A ist dagegen eine Abbildung F: A ® [0,1]. Der Wert f(x) kann als Grad der Zugehörigkeit gedeutet werden f(x) =1 volle Zugehörigkeit, f(x) = 0 keine Zugehörigkeit. Die Klasse der reellen Zahlen, die viel größer sind als 1, kann beispielsweise durch die fuzzy-Menge dargestellt werden (vgl. Diagramm oben rechts): f(x) = 0 für x = 1, 1-exp(1-x) für x >1.
phorylierte Zustand einer digitalen "1" und der nichtaktive, unphosphorylierte einer digitalen "0". Wenn mehrere Proteine kooperieren, kann zwischen 0 und 1 wie bei einem Ein-Aus-Schalter hin- und hergeschaltet werden.
Proteine, die mehrere Bindungsstellen besitzen, sind keine Seltenheit. Solche Proteine können logische Verknüpfungen vornehmen. Sie sind beispielsweise nur dann aktiv, wenn gleichzeitig an zwei Stellen Phosphate sitzen, entsprechend einer logischen UND-Verknüpfung. Oder sie sind aktiv, wenn sie an mindestens einer Stelle phosphoryliert sind, was einer logischen ODER-Verknüpfung entspricht.
Darüber hinaus ist in manchen Proteinen auch eine "unscharfe Logik", eine fuzzy-Logik, realisiert, bei der es Zwischenzustände gibt. Solche fuzzy-Proteine können nicht nur Entweder-Oder, sondern auch Informationen wie "etwas wärmer" oder "etwas dunkler" verarbeiten.
Chemische Netzwerke sind wesentlich komplexer als die Netzwerke elektronischer Computer. Im Gegensatz zum Computer kennt eine Zelle keine Elektronenströme und keine Kabel. In chemischen Netzwerken werden Signale entweder durch direkten Kontakt zwischen Proteinen oder durch Botenstoffe - beispielsweise phosphorylierte Moleküle - übermittelt. Die Botenstoffe treiben frei in der Zelle, sie "diffundieren", bis sie an Proteine geraten, die die Botschaft verstehen und umsetzen.
Eine einzige biologische Zelle besitzt eine millionenfach kleinere Fläche als ein Mikrochip - dennoch beherbergt sie Hunderte verschiedener Rezeptoren und Proteine, die sich gegenseitig beeinflussen. In der biologischen Zelle ist, anders als im Computer und noch extremer als bei den Nervenzellen im Gehirn, jede Einheit mit jeder anderen verbunden.
Die Effekte auf die Aktivität und Konzentration von Tausenden von weiteren Zellproteinen, die für den Stoffwechsel und die Bewegung der Zelle zuständig sind, sind oft hochgradig nichtlinear und mathematisch kaum zu beschreiben. Im Extremfall treten Signalverstärkungen um den Faktor 100 000 auf: So viele Moleküle kann beispielsweise ein einzelnes Lichtteilchen verändern, das auf einen Photorezeptor fällt.
Trotz dieser hohen Vernetzung und des unspezifischen Signaltransportes ist die Geschwindigkeit der Informationsübermittlung erstaunlich hoch: Binnen einer Zehntelsekunde erreichen die diffundierenden Botenmoleküle jeden Ort in der Zelle.
Zellsysteme speichern Wissen als Verknüpfungsmuster biochemischer oder struktureller Veränderungen. So spiegelt beispielsweise die Konzentration der phosphorylierten Enzyme im Zellinneren genau die chemische Umgebung wider, in der sich die Zelle innerhalb der letzten Sekunden befunden hat.
Dagegen haben die Inhalte der Speicherzellen eines Rechners zueinander keinerlei Beziehung. Jedes Bit steht im Speicher für sich, unabhängig von anderen gespeicherten Bits. Die gegenseitige Unabhängigkeit der Bits bleibt selbst dann erhalten, wenn Informationen im Rechner aus dem Speicher abgerufen und durch elektrische Größen dargestellt werden.
2.3.3 Das Lösen von Problemen
Schon einzellige Organismen verfügen über die Fähigkeit, eigene Lösungen zu einem besonderen Problem zu entwickeln. Sie brauchen keinen wohldefinierten Prozeß, mit dessen Hilfe sie eine Eingabe in eine Ausgabe überführen. Regeln werden nicht explizit gelernt, sondern implizit generalisiert, wobei sich biologische Systeme veränderten Regeln selbständig anpassen können.
Ein Computer hingegen führt fest vorgegebene Handlungsschritte aus - er selbst entwickelt keine Lösung zu einem Problem. Im Gegensatz zu natürlichen Systemen kennt er nur eine einzige Art von Daten, nämlich binäre Ziffern, und er arbeitet stets nach dem gleichen Satz von Regeln, nämlich dem eines ihm vorgegebenen Befehlssatzes, bestehend aus einzelnen Mikrooperationen. Ihre Ausführung bedeutet die Umwandlung einer Information in ein elektrisches Signal, das einen Schaltvorgang im Rechner auslöst. Zahlreiche Mikroinstruktionen werden vom Rechner automatisch hintereinander ausgeführt und bilden einen Maschinenbefehl - etwas, das einem Programmierer als kleinste unteilbare Einheit erscheint. Die einen Maschinenbefehl ausführende Folge von Mikrobefehlen bildet ein Mikroprogramm.
Die Art der Mikroprogrammierung entscheidet über die Flexibilität eines Rechners. So können Mikroprogramme fest verdrahtet oder in besonderen Speichern abgelegt sein. Für diese Speicher verwendet man gewöhnlich zwei Arten: einen Nur-Lese-Speicher, abgekürzt ROM (read only memory), und einen Schreib-Lese-Speicher mit wahlfreiem Zugriff (RAM, random access memory). Der Nur-Lese-Speicher enthält Informationen, die vom Hersteller fest einprogrammiert wurden und sich nicht mehr verändern lassen. Bei einem Schreib-Lese-Speicher können Informationen dagegen jederzeit nach Bedarf eingeschrieben und gelesen werden. Entspricht das Proteinnetzwerk natürlicher Zellen dem "random access memory", so kann die in den Erbanlagen der DNA niedergelegte genetische Information mit dem Dauerspeicher und Betriebssystem eines Computers verglichen werden.
Es ergibt sich eine Schwierigkeit: Die Probleme, die ein Computer lösen soll, sind kaum jemals in seiner Sprache, d.h. mit Hilfe von Bits, formuliert. Statt dessen ist der Lösungsweg eines Problems von einem Menschen in Form eines Algorithmus beschrieben, eines aus endlich vielen Schritten bestehenden Verarbeitungsverfahrens. Die zu verarbeitenden Daten sind als Zahlen, Schriftzeichen, Symbole usw. ausgedrückt. Der Algorithmus selbst ist in natürlicher Sprache abgefaßt, die voller Ungenauigkeiten und Mehrdeutigkeiten ist. Oft können diese nur durch Hintergrundwissen aufgelöst werden - etwas, über das ein Computer nicht verfügt. Zudem faßt die natürliche Sprache viele Einzelvorgänge zu einem Ganzen zusammen bzw. setzt sie implizit in Beziehung zueinander. Darin ähnelt sie der Informationsverarbeitung biologischer Zellsysteme, die gleichzeitig auf genomischer, molekularer, auf System- und Verhaltensebene stattfindet. Wie die natürliche Sprache verarbeiten biologische Systeme auch Ungenaues (etwas, das "fuzzy" ist).
Ein Computer der heutigen Rechnergeneration hingegen akzeptiert nur einen strengen Formalismus. Er ist nicht mehr als eine Maschine, die physikalische Regeln in ihm vorgegebener Form manipuliert. Zwar wäre es theoretisch möglich, ein fuzzy-System auf einem fuzzy-Computer zu verwirklichen, wie es von Zadeh angeregt wurde, doch diese Möglichkeit wurde bisher nicht verwirklicht. Bis zur Herstellung eines fuzzy-Computers bleibt nur die Simulation mit Hilfe von mathematischen Formeln, die, übersetzt in die Rechnersprache, von einem Computer abgearbeitet werden können.
So muß eine Vielfalt von Informationsdarstellungen von einer Maschine mit nur einem einfachen Sprachschatz - einem einzigen, festen Befehlssatz - verstanden werden. Hierzu wird der in natürlicher Sprache formulierte Algorithmus zunächst in eine Sprache übersetzt, in der die Bedeutung jedes Objektes und jeder Anweisung präzise und eindeutig definiert ist - in eine Programmiersprache. In ihr sind die Objekte des Algorithmus in genau festgelegter Form als Daten repräsentiert. Die Objekte erhalten Namen, genannt Variablen, wenn sie veränderlich sind, und Konstanten, wenn sie gleich bleiben.
Zusammen mit den Daten enthält das Programm eine vollständige Anweisungsfolge, die angibt, auf welche Art die Daten verändert werden sollen. Eine Programmiersprache ist nach strengen Regeln aufgebaut. Dennoch hat sie eine oberflächliche Ähnlichkeit mit einer natürlichen Sprache. Sie verfügt über ein Vokabular von Wörtern, Zahlen und Schlüsselwörtern mit einer festen Bedeutung. Für die Kombination der Grundsymbole existieren Regeln, die eine Grammatik bilden.
Es gibt zwei Arten von Programmiersprachen. Während die problemorientierten Sprachen von der jeweiligen Computeranlage unabhängig sind, nutzen maschinenorientierte Sprachen die technischen Eigenschaften einer Anlage besonders gut aus, sind aber anlagenabhängig.
Der Schritt von der Programmiersprache in die Sprache des Rechners, die Maschinensprache, - wird vom Computer selbst vor-
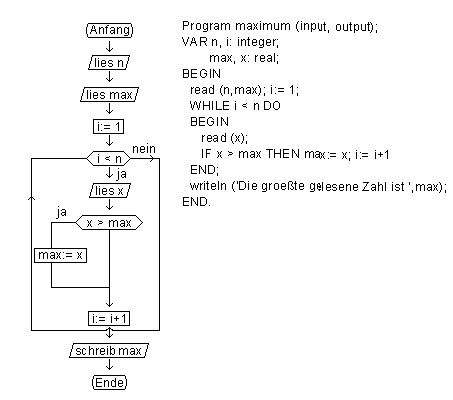
Bild 2.19
An dem Problem, die größte unter n natürlichen Zahlen zu bestimmen, sollen die Besonderheiten einer Programmiersprache gezeigt werden. Links ist das Ablaufprogramm eines Algorithmus für das Problem dargestellt. Die Übersetzung des Algorithmus in die Programmiersprache Pascal, eine problemorientierte Sprache, ist rechts gezeigt. Kernstück des Algorithmus ist eine n-mal auszuführende Schleife. Bei jedem Durchlaufen der Schleife wird eine Zahl x eingelesen. Ist sie größer als die größte der bisher eingelesenen Zahlen (genannt "max"), nimmt max den Wert von x an.genommen. Überführen in biologischen Systemen Moleküle Information von einer Form in die andere, so wandeln in Computern Übersetzungsprogramme (Compiler bzw. Assembler genannt) problemorientierte bzw. maschinenorientierte Sprachen in Maschinensprachen um. Da die Hardware eines Computers nur die elektronische Darstellung binärer Zahlen kennt, besteht die Maschinensprache aus einer Folge solcher Zahlen.
Diese Zahlenfolge bildet Befehlsworte. Sie enthalten einen Operationsteil, der bestimmt, welche Operationen der Rechner vornehmen soll, und einen Adressenteil mit den Adressen der Operanden, die der Rechner bei dieser Operation anzusprechen hat. Die im Rechner gespeicherte Adresse ähnelt dem Rezeptor der natürlichen Zelle - auch er dient wie die Adresse dazu, ein Ding an seinen richtigen Wirkort zu führen.
|
Operationsteil |
Adreßteil |
Adreßteil |
|
ADD |
143 |
298 |
|
Was |
Woher/Wohin |
Woher |
Bild 2.20
Beispiel einer Addition zweier Zahlen in Maschinensprache. In unsere Sprache übersetzt, könnte der Maschinenbefehl so ausgedrückt werden: Hole die erste Zahl, die im Arbeitsspeicher unter der Adresse 143 steht, in das Rechenwerk. Addiere zu ihr die zweite Zahl, die unter der Adresse 298 gespeichert ist. Bringe das Ergebnis nach Speicherstelle 143.2.3.4 Die Zeit
Die Ausführung eines Mikroprogramms geschieht nach einem vorgegebenen Zeitraster, d.h. einer Folge von Taktintervallen. Innerhalb eines Taktintervalls werden verschiedene Mikrooperationen gleichzeitig angestoßen. Sie alle zusammen bilden den Mikrobefehl.
Ein Taktgenerator erzeugt eine gleichmäßige Impulsfolge, die die gesamte Befehlsfolge des Mikroprogramms zeitlich synchronisiert. Wichtigster Bestandteil des Taktgenerators sind kristalline Quarze, deren gleichmäßige Schwingungen Grundlage einer regelmäßigen Taktfrequenz sind.
|
Abweichung vom Mittelwert in % |
|
Uhrzeit |
Bild 2.21
Änderung der Cortisolkonzentration im Blut im Verlaufe eines TagesEbenso wie die Ausführung eines Mikroprogramms sind alle Lebensvorgänge einer Zelle zeitlich organisiert. Im Unterschied zum Mikroprogramm sind die Zitmuster im Zellverhalten jedoch komplex. Nicht nur Vorgänge der inneren, sondern auch der äußeren Umwelt werden als Zeitgeber wirksam, an dem sich das Zeitmuster einer Zelle orientiert. So können beispielsweise äußere Perioden wie Tagesrhythmen und Jahresrhythmen als Zeitgeber wirksam sein. Einfache Zeitgeber, d.h. ähnlich den Quarzkristallen schwingendeSysteme, haben Einfluß auf die Geschwindigkeit, mit der Moleküle in biologischen Systemen Informationen verarbeiten.
Zu den tagesperiodischen Schwankungen gehören die circadianen Schwankungen der Kernvolumina, die mit rhythmischen Synthesevorgängen (Nukleotidsynthese) zusammenhängen. Auch das Zytoplasma bestimmter Zellen kann tägliche Volumenschwankungen aufweisen. Dies ist beispielsweise bei Zellen der Nebennierenrinde der Fall, die das Hormon Cortisol produzieren, dessen Konzentration im Blut einen charakteristischen circadianen Rhythmus zeigt.
Die Beispiele zeigen: Im Gegensatz zum Computer gibt es keine einheitliche biologische Zeit, aber biologische Zeitstandards, die einer unaufhörlichen Dynamik unterworfen sind.